
Food web facts for kids

A food web is similar to a food chain but larger,and it is a diagram that combines many food chains into one picture.
Food webs show how plants and animals are connected in many ways to help them all survive, unlike food chains that just follow one path.
For example, a tree produces acorns - this is called the producer. Mice, squirrels, and insects eat these acorns and because there are many mice, the weasels, snakes and raccoons have food. With insects in the acorns, other predators would be attracted (like skunks or opossums) and therefore, foxes, hawks or owls could all find food. Food webs can be effected in many ways considering the urban areas of major countries are taking no action and care. It uses arrows to show the energy relationships among organisms.
Charles Elton pioneered the concept of food cycles, food chains, and food size in his classical 1927 book "Animal Ecology"; Elton's 'food cycle' was replaced by 'food web' in a subsequent ecological text. Elton organized species into functional groups, which was the basis for Raymond Lindeman's classic and landmark paper in 1942 on trophic dynamics. Lindeman emphasized the important role of decomposer organisms in a trophic system of classification. The notion of a food web has a historical foothold in the writings of Charles Darwin and his terminology, including an "entangled bank", "web of life", "web of complex relations", and in reference to the decomposition actions of earthworms he talked about "the continued movement of the particles of earth". Even earlier, in 1768 John Bruckner described nature as "one continued web of life".
Food webs are limited representations of real ecosystems as they necessarily aggregate many species into trophic species, which are functional groups of species that have the same predators and prey in a food web. Ecologists use these simplifications in quantitative (or mathematical representation) models of trophic or consumer-resource systems dynamics. Using these models they can measure and test for generalized patterns in the structure of real food web networks. Ecologists have identified non-random properties in the topological structure of food webs. Published examples that are used in meta analysis are of variable quality with omissions. However, the number of empirical studies on community webs is on the rise and the mathematical treatment of food webs using network theory had identified patterns that are common to all. Scaling laws, for example, predict a relationship between the topology of food web predator-prey linkages and levels of species richness.
History of food webs

Food webs serve as a framework to help ecologists organize the complex network of interactions among species observed in nature and around the world. One of the earliest descriptions of a food chain was described by a medieval Afro-Arab scholar named Al-Jahiz: "All animals, in short, cannot exist without food, neither can the hunting animal escape being hunted in his turn." The earliest graphical depiction of a food web was by Lorenzo Camerano in 1880, followed independently by those of Pierce and colleagues in 1912 and Victor Shelford in 1913. Two food webs about herring were produced by Victor Summerhayes and Charles Elton and Alister Hardy in 1923 and 1924. Charles Elton subsequently pioneered the concept of food cycles, food chains, and food size in his classical 1927 book "Animal Ecology"; Elton's 'food cycle' was replaced by 'food web' in a subsequent ecological text. After Charles Elton's use of food webs in his 1927 synthesis, they became a central concept in the field of ecology. Elton organized species into functional groups, which formed the basis for the trophic system of classification in Raymond Lindeman's classic and landmark paper in 1942 on trophic dynamics. The notion of a food web has a historical foothold in the writings of Charles Darwin and his terminology, including an "entangled bank", "web of life", "web of complex relations", and in reference to the decomposition actions of earthworms he talked about "the continued movement of the particles of earth". Even earlier, in 1768 John Bruckner described nature as "one continued web of life".
Interest in food webs increased after Robert Paine's experimental and descriptive study of intertidal shores suggesting that food web complexity was key to maintaining species diversity and ecological stability. Many theoretical ecologists, including Sir Robert May and Stuart Pimm, were prompted by this discovery and others to examine the mathematical properties of food webs.
Kinds of food webs
Food webs are necessarily aggregated and only illustrate a tiny portion of the complexity of real ecosystems. For example, the number of species on the planet are likely in the general order of 107, over 95% of these species consist of microbes and invertebrates, and relatively few have been named or classified by taxonomists. It is explicitly understood that natural systems are 'sloppy' and that food web trophic positions simplify the complexity of real systems that sometimes overemphasize many rare interactions. Most studies focus on the larger influences where the bulk of energy transfer occurs. "These omissions and problems are causes for concern, but on present evidence do not present insurmountable difficulties."

There are different kinds or categories of food webs:
- Source web - one or more node(s), all of their predators, all the food these predators eat, and so on.
- Sink web - one or more node(s), all of their prey, all the food that these prey eat, and so on.
- Community (or connectedness) web - a group of nodes and all the connections of who eats whom.
- Energy flow web - quantified fluxes of energy between nodes along links between a resource and a consumer.
- Paleoecological web - a web that reconstructs ecosystems from the fossil record.
- Functional web - emphasizes the functional significance of certain connections having strong interaction strength and greater bearing on community organization, more so than energy flow pathways. Functional webs have compartments, which are sub-groups in the larger network where there are different densities and strengths of interaction. Functional webs emphasize that "the importance of each population in maintaining the integrity of a community is reflected in its influence on the growth rates of other populations."
Within these categories, food webs can be further organized according to the different kinds of ecosystems being investigated. For example, human food webs, agricultural food webs, detrital food webs, marine food webs, aquatic food webs, soil food webs, Arctic (or polar) food webs, terrestrial food webs, and microbial food webs. These characterizations stem from the ecosystem concept, which assumes that the phenomena under investigation (interactions and feedback loops) are sufficient to explain patterns within boundaries, such as the edge of a forest, an island, a shoreline, or some other pronounced physical characteristic.
Detrital web
In a detrital web, plant and animal matter is broken down by decomposers, e.g., bacteria and fungi, and moves to detritivores and then carnivores. There are often relationships between the detrital web and the grazing web. Mushrooms produced by decomposers in the detrital web become a food source for deer, squirrels, and mice in the grazing web. Earthworms eaten by robins are detritivores consuming decaying leaves.
Images for kids
-
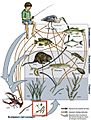
A freshwater aquatic food web. The blue arrows show a complete food chain (algae → daphnia → gizzard shad → largemouth bass → great blue heron)
-
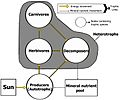
A simplified food web illustrating a three trophic food chain (producers-herbivores-carnivores) linked to decomposers. The movement of mineral nutrients is cyclic, whereas the movement of energy is unidirectional and noncyclic. Trophic species are encircled as nodes and arrows depict the links.
-
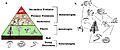
A trophic pyramid (a) and a simplified community food web (b) illustrating ecological relations among creatures that are typical of a northern Boreal terrestrial ecosystem. The trophic pyramid roughly represents the biomass (usually measured as total dry-weight) at each level. Plants generally have the greatest biomass. Names of trophic categories are shown to the right of the pyramid. Some ecosystems, such as many wetlands, do not organize as a strict pyramid, because aquatic plants are not as productive as long-lived terrestrial plants such as trees. Ecological trophic pyramids are typically one of three kinds: 1) pyramid of numbers, 2) pyramid of biomass, or 3) pyramid of energy.
-

Multitrophic interaction: Euphydryas editha taylori larvae sequester defensive compounds from specific types of plants they consume to protect themselves from bird predators
-
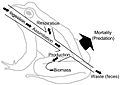
Energy flow diagram of a frog. The frog represents a node in an extended food web. The energy ingested is utilized for metabolic processes and transformed into biomass. The energy flow continues on its path if the frog is ingested by predators, parasites, or as a decaying carcass in soil. This energy flow diagram illustrates how energy is lost as it fuels the metabolic process that transform the energy and nutrients into biomass.
-
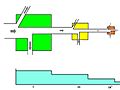
An expanded three link energy food chain (1. plants, 2. herbivores, 3. carnivores) illustrating the relationship between food flow diagrams and energy transformity. The transformity of energy becomes degraded, dispersed, and diminished from higher quality to lesser quantity as the energy within a food chain flows from one trophic species into another. Abbreviations: I=input, A=assimilation, R=respiration, NU=not utilized, P=production, B=biomass.
-
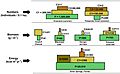
Illustration of a range of ecological pyramids, including top pyramid of numbers, middle pyramid of biomass, and bottom pyramid of energy. The terrestrial forest (summer) and the English Channel ecosystems exhibit inverted pyramids.Note: trophic levels are not drawn to scale and the pyramid of numbers excludes microorganisms and soil animals. Abbreviations: P=Producers, C1=Primary consumers, C2=Secondary consumers, C3=Tertiary consumers, S=Saprotrophs.
-
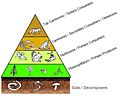
A four level trophic pyramid sitting on a layer of soil and its community of decomposers.
-
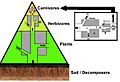
A three layer trophic pyramid linked to the biomass and energy flow concepts.
-
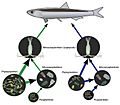
A simplified version of a food web in the Gulf of Naples in eutrophic (Green) and oligotrophic (Blue) summer conditions. In the Green system state, both copepods and microzooplankton exert a strong grazing pressure on phytoplankton, while in the Blue state, copepods increase their predation over microzooplankton, which in turn shifts its predation from phytoplankton to bacterial plankton or picoplankton. These trophic mechanisms stabilize the delivery of organic matter from copepods to fish.










See also
 In Spanish: Red trófica para niños
In Spanish: Red trófica para niños
