
Edmontosaurus facts for kids
Quick facts for kids EdmontosaurusTemporal range: Upper Cretaceous 73–65.5 mya
|
|
|---|---|
 |
|
| Mounted skeleton of E. regalis, Oxford University Museum of Natural History | |
| Scientific classification | |
| Kingdom: | |
| Class: | |
| Superorder: | |
| Order: | |
| Suborder: | |
| Family: | |
| Genus: |
Edmontosaurus
|
Edmontosaurus was a large, plant-eating, duck-billed dinosaur from the latest Cretaceous stage. It had short arms, a long, pointed tail, three-toed, hoofed feet, and mitten-like hands. Its head was flat and sloping with a wide, toothless beak, cheek pouches, and hundreds of closely-packed cheek teeth that ground up its food. It is the same animal as Anatotitan. There are two species, and the animal has been called by several other names in the past.
Edmontosaurus was named from a fossil found in the Horseshoe Canyon Formation, in Alberta, Canada. Many Edmontosaurus fossils have been found, in Alberta, and Alaska, Wyoming, Montana and New Jersey in the USA. Two mummified Edmontosaurus fossils were found in Wyoming.
The distribution of its fossils suggests that it preferred coasts and coastal plains. It could move on two legs and on four. Because it is known from several bone beds, Edmontosaurus is thought to have lived in groups. It may have been migratory.
Contents
Description

Edmontosaurus has been described in detail from numerous specimens. Like other hadrosaurids, it was a bulky animal with a long, laterally flattened tail and a head with an expanded, duck-like beak. The skull had no bony crest, unlike many other hadrosaurids, but bore a comb-like crest of skin and scales. The fore legs were not as heavily built as the hind legs, but were long enough to be used in standing or movement. Edmontosaurus was among the largest hadrosaurids: depending on the species, a fully grown adult could have been 9 metres (30 ft) long, and some of the larger specimens reached the range of 12 metres (39 ft) to 13 metres (43 ft) long. Its weight was on the order of 4.0 metric tons (4.4 short tons). Traditionally, E. regalis has been regarded as the largest species, though this was challenged by the hypothesis that the larger hadrosaurid Anatotitan copei is a synonym of Edmontosaurus annectens, as put forward by Jack Horner and colleagues in 2004, and supported in studies by Campione and Evens in 2009 and 2011. The type specimen of E. regalis, NMC 2288, is estimated as 9 to 12 metres (30 to 39 ft) long. E. annectens is often seen as smaller. Two mounted skeletons, USNM 2414 and YPM 2182, measure 8.00 metres (26.25 ft) long and 8.92 metres (29.3 ft) long, respectively. However, these are probably subadult individuals, and there is at least one report of a much larger potential E. annectens specimen, almost 12 metres (39 ft) long. Two specimens still under study in the collection of the Museum of the Rockies - a 7.6 m (25 ft) tail labelled as MOR 1142 and another labelled as MOR 1609 - indicate that Edmontosaurus annectens could have grown to larger sizes, possibly rivaling Shantungosaurus in size. The specimens of these individuals indicate a length of up to 15 m (49 ft), according to paleontologists. Such large individuals of Edmontosaurus would likely have been very rare due to such factors as environmental stress, disease, and predation.
Skull

The skull of a fully grown Edmontosaurus could be over a metre long. One skull of E. annectens (formerly Anatotitan) measures 3.87 feet (1.18 m) long. The skull was roughly triangular in profile, with no bony cranial crest. Viewed from above, the front and rear of the skull were expanded, with the broad front forming a duck-bill or spoon-bill shape. The beak was toothless, and both the upper and lower beaks were extended by keratinous material. Substantial remains of the keratinous upper beak are known from the "mummy" kept at the Senckenberg Museum. In this specimen, the preserved nonbony part of the beak extended for at least 8 centimetres (3.1 in) beyond the bone, projecting down vertically. The nasal openings of Edmontosaurus were elongate and housed in deep depressions surrounded by distinct bony rims above, behind, and below. In at least one case (the Senckenberg specimen), rarely preserved sclerotic rings were preserved in the eye sockets. Another rarely seen bone, the stapes (the reptilian ear bone), has also been seen in a specimen of Edmontosaurus.
Teeth were present only in the maxillae (upper cheeks) and dentaries (main bone of the lower jaw). The teeth were continually replaced, taking about half a year to form. They were composed of six types of tissues, rivaling the complexity of mammal teeth. They grew in columns, with an observed maximum of six in each, and the number of columns varied based on the animal's size. Known column counts for the two species are: 51 to 53 columns per maxilla and 48 to 49 per dentary (teeth of the upper jaw being slightly narrower than those in the lower jaw) for E. regalis; and 52 columns per maxilla and 44 per dentary for E. annectens (an E. saskatchewanensis specimen).
Postcranial skeleton

The number of vertebrae differs between specimens. E. regalis had thirteen neck vertebrae, eighteen back vertebrae, nine hip vertebrae, and an unknown number of tail vertebrae. A specimen once identified as belonging to Anatosaurus edmontoni (now considered to be the same as E. regalis) is reported as having an additional back vertebra and 85 tail vertebrae, with an undisclosed amount of restoration. Other hadrosaurids are only reported as having 50 to 70 tail vertebrae, so this appears to have been an overestimate. The anterior back was curved toward the ground, with the neck flexed upward and the rest of the back and tail held horizontally. Most of the back and tail were lined by ossified tendons arranged in a latticework along the neural spines of the vertebrae. This condition has been described as making the back and at least part of the tail "ramrod" straight. The ossified tendons are interpreted as having strengthened the vertebral column against gravitational stress, incurred through being a large animal with a horizontal vertebral column otherwise supported mostly by the hind legs and hips.

The shoulder blades were long flat blade-like bones, held roughly parallel to the vertebral column. The hips were composed of three elements each: an elongate ilium above the articulation with the leg, an ischium below and behind with a long thin rod, and a pubis in front that flared into a plate-like structure. The structure of the hip hindered the animal from standing with its back erect, because in such a position the thigh bone would have pushed against the joint of the ilium and pubis, instead of pushing only against the solid ilium. The nine fused hip vertebrae provided support for the hip.
The fore legs were shorter and less heavily built than the hind legs. The upper arm had a large deltopectoral crest for muscle attachment, while the ulna and radius were slim. The upper arm and forearm were similar in length. The wrist was simple, with only two small bones. Each hand had four fingers, with no thumb (first finger). The index second, third, and fourth fingers were approximately the same length and were united in life within a fleshy covering. Although the second and third finger had hoof-like unguals, these bones were also within the skin and not apparent from the outside. The little finger diverged from the other three and was much shorter. The thigh bone was robust and straight, with a prominent flange about halfway down the posterior side. This ridge was for the attachment of powerful muscles attached to the hips and tail that pulled the thighs (and thus the hind legs) backward and helped maintain the use of the tail as a balancing organ. Each foot had three toes, with no big toe or little toe. The toes had hoof-like tips.
Skin

Multiple specimens of Edmontosaurus annectens have been found with preserved skin impressions. Several have been well-publicized, such as the "Trachodon mummy" of the early 20th century, and the specimen nicknamed "Dakota", the latter apparently including remnant organic compounds from the skin. Because of these finds, the scalation of Edmontosaurus annectens is known for most areas of the body. Skin impressions are less well known for E. regalis, but some well-preserved examples have been studied, including one which preserves a soft tissue crest or wattle on the head. It is unknown whether such a crest was present on E. annectens, and whether it was an indicator of sexual dimorphism.
Discovery and history
Claosaurus annectens

The first well-supported species of Edmontosaurus was named in 1892 as Claosaurus annectens by Othniel Charles Marsh. This species is based on USNM 2414, a partial skull-roof and skeleton, with a second skull and skeleton, YPM 2182, designated the paratype. Both were collected in 1891 by John Bell Hatcher from the late Maastrichtian-age Upper Cretaceous Lance Formation of Niobrara County (then part of Converse County), Wyoming. This species has some historical footnotes attached: it is among the first dinosaurs to receive a skeletal restoration, and is the first hadrosaurid so restored; and YPM 2182 and UNSM 2414 are, respectively, the first and second essentially complete mounted dinosaur skeletons in the United States. YPM 2182 was put on display in 1901, and USNM 2414 in 1904.
Charles W. Gilmore in 1915 reassessed hadrosaurids and recommended that Thespesius be reintroduced for hadrosaurids from the Lance Formation and rock units of equivalent age, and that Trachodon, based on inadequate material, should be restricted to a hadrosaurid from the older Judith River Formation and its equivalents. In regards to Claosaurus annectens, he recommended that it be considered the same as Thespesius occidentalis. His reinstatement of Thespesius for Lance-age hadrosaurids would have other consequences for the taxonomy of Edmontosaurus in the following decades.
During this time frame (1902–1915), two additional important specimens of C. annectens were recovered. The first, the "Trachodon mummy" (AMNH 5060), was discovered in 1908 by Charles Hazelius Sternberg and his sons in Lance Formation rocks near Lusk, Wyoming. Sternberg was working for the British Museum of Natural History, but Henry Fairfield Osborn of the American Museum of Natural History was able to purchase the specimen for $2,000.
Canadian discoveries
Edmontosaurus itself was coined in 1917 by Lawrence Lambe for two partial skeletons found in the Horseshoe Canyon Formation (formerly the lower Edmonton Formation) along the Red Deer River of southern Alberta, Canada. These rocks are older than the rocks in which Claosaurus annectens was found. The Edmonton Formation lends Edmontosaurus its name. The type species, E. regalis ("regal", or, more loosely, "king-sized"), is based on NMC 2288, consisting of a skull, articulated vertebrae up to the sixth tail vertebra, ribs, partial hips, an upper arm bone, and most of a hind limb. It was discovered in 1912 by Levi Sternberg. The second specimen, paratype NMC 2289, consists of a skull and skeleton lacking the beak, most of the tail, and part of the feet. It was discovered in 1916 by George F. Sternberg. Lambe found that his new dinosaur compared best to Diclonius mirabilis (specimens now assigned to Edmontosaurus annectens), and drew attention to the size and robustness of Edmontosaurus. Initially, Lambe only described the skulls of the two skeletons, but returned to the genus in 1920 to describe the skeleton of NMC 2289. The postcrania of the type specimen remains undescribed, still in its plaster jackets.

Two more species that would come to be included with Edmontosaurus were named from Canadian remains in the 1920s, but both would initially be assigned to Thespesius. Gilmore named the first, Thespesius edmontoni, in 1924. T. edmontoni also came from the Horseshoe Canyon Formation. It was based on NMC 8399, another nearly complete skeleton lacking most of the tail. NMC 8399 was discovered on the Red Deer River in 1912 by a Sternberg party. Its forelimbs, ossified tendons, and skin impressions were briefly described in 1913 and 1914 by Lambe, who at first thought it was an example of a species he had named Trachodon marginatus, but then changed his mind. The specimen became the first dinosaur skeleton to be mounted for exhibition in a Canadian museum. Gilmore found that his new species compared closely to what he called Thespesius annectens, but left the two apart because of details of the arms and hands. He also noted that his species had more vertebrae than Marsh's in the back and neck, but proposed that Marsh was mistaken in assuming that the annectens specimens were complete in those regions.
Species and distribution

Edmontosaurus is currently regarded as having two valid species: type species E. regalis, and E. annectens. E. regalis is known only from the Horseshoe Canyon Formation of Alberta, dating from the late Campanian stage of the late Cretaceous period. At least a dozen individuals are known, including seven skulls with associated postcrania, and five to seven other skulls. The species formerly known as Thespesius edmontoni or Anatosaurus edmontoni represents immature individuals.
E. annectens is known from the Frenchman Formation of Saskatchewan, the Hell Creek Formation of Montana, and the Lance Formation of South Dakota and Wyoming. It is limited to late Maastrichtian rocks, and is represented by at least twenty skulls, some with postcranial remains. One author, Kraig Derstler, has described E. annectens as "perhaps the most perfectly-known dinosaur to date [1994]." Anatosaurus copei and E. saskatchewanensis are now thought to be growth stages of E. annectens: A. copei as adults, and E. saskatchewanensis as juveniles. Trachodon longiceps may be a synonym of E. annectens as well. Anatosaurus edmontoni was mistakenly listed as a synonym of E. annectens in both reviews of the Dinosauria, but this does not appear to be the case.

E. annectens differed from E. regalis by having a longer, lower, less robust skull. Although Brett-Surman regarded E. regalis and E. annectens as potentially representing males and females of the same species, all E. regalis specimens come from older formations than E. annectens specimens.
Edmontosaurin specimens from the Prince Creek Formation of Alaska formerly assigned to Edmontosaurus sp. have been given their own genus and species name, Ugrunaaluk kuukpikensis. However, the identification of Ugrunaaluk as a separate genus was questioned by a 2017 study from Hai Xing and colleagues, who regarded it as a nomen dubium indistinguishable from other Edmontosaurus. Edmontosaurus was also reported from the Javelina Formation of Big Bend National Park, western Texas based on TMM 41442-1, but was later referred to Kritosaurus cf. navajovius by Wagner (2001), before being assigned to Kritosaurus sp. by Lehman et al. (2016).
Paleobiology
Growth

In a 2011 study, Campione and Evans recorded data from all known "edmontosaur" skulls from the Campanian and Maastrichtian and used it to plot a morphometric graph, comparing variable features of the skull with skull size. Their results showed that within both recognized Edmontosaurus species, many features previously used to classify additional species or genera were directly correlated with skull size. Campione and Evans interpreted these results as strongly suggesting that the shape of Edmontosaurus skulls changed dramatically as they grew. This has led to several apparent mistakes in classification in the past. The Campanian species Thespesius edmontoni, previously considered a synonym of E. annectens due to its small size and skull shape, is more likely a subadult specimen of the contemporary E. regalis. Similarly, the three previously recognized Maastrichtian edmontosaur species likely represent growth stages of a single species, with E. saskatchewanensis representing juveniles, E. annectens subadults, and Anatotitan copei fully mature adults. The skulls became longer and flatter as the animals grew.
Diet
Feeding adaptations

As a hadrosaurid, Edmontosaurus was a large terrestrial herbivore. Its teeth were continually replaced and packed into dental batteries that contained hundreds of teeth, only a relative handful of which were in use at any time. It used its broad beak to cut loose food, perhaps by cropping, or by closing the jaws in a clamshell-like manner over twigs and branches and then stripping off the more nutritious leaves and shoots. Because the tooth rows are deeply indented from the outside of the jaws, and because of other anatomical details, it is inferred that Edmontosaurus and most other ornithischians had cheek-like structures, muscular or non-muscular. The function of the cheeks was to retain food in the mouth. The animal's feeding range would have been from ground level to around 4 metres (13 ft) above.

A study published in 2008 by Casey Holliday and Lawrence Witmer found that ornithopods like Edmontosaurus lacked the types of skull joints seen in those modern animals that are known to have kinetic skulls (skulls that permit motion between their constituent bones), such as squamates and birds. They proposed that joints that had been interpreted as permitting movement in dinosaur skulls were actually cartilaginous growth zones. An important piece of evidence for Weishampel's model is the orientation of scratches on the teeth, showing the direction of jaw action. Other movements could produce similar scratches though, such as movement of the bones of the two halves of the lower jaw. Not all models have been scrutinized under present techniques. Vincent Williams and colleagues (2009) published additional work on hadrosaurid tooth microwear. They found four classes of scratches on Edmontosaurus teeth. The most common class was interpreted as resulting from an oblique motion, not a simple up-down or front-back motion, which is consistent with the Weishampel model. This motion is thought to have been the primary motion for grinding food. Two scratch classes were interpreted as resulting from forward or backward movement of the jaws. The other class was variable and probably resulted from opening the jaws. The combination of movements is more complex than had been previously predicted.

Weishampel developed his model with the aid of a computer simulation. Natalia Rybczynski and colleagues have updated this work with a much more sophisticated three-dimensional animation model, scanning a skull of E. regalis with lasers. They were able to replicate the proposed motion with their model, although they found that additional secondary movements between other bones were required, with maximum separations of 1.3 to 1.4 centimetres (0.51 to 0.55 in) between some bones during the chewing cycle. Rybczynski and colleagues were not convinced that the Weishampel model is viable, but noted that they have several improvements to implement to their animation. Planned improvements include incorporating soft tissue and tooth wear marks and scratches, which should better constrain movements. They note that there are several other hypotheses to test as well. Further research published in 2012 by Robin Cuthbertson and colleagues found the motions required for Weishampel's model to be unlikely, and favored a model in which movements of the lower jaw produced grinding action. The lower jaw's joint with the upper jaw would permit anterior–posterior motion along with the usual rotation, and the anterior joint of the two halves of the lower jaw would also permit motion; in combination, the two halves of the lower jaw could move slightly back and forth as well as rotating slightly along their long axes. These motions would account for the observed tooth wear and a more solidly constructed skull than modeled by Weishampel.
Because scratches dominate the microwear texture of the teeth, Williams et al. suggested Edmontosaurus was a grazer instead of a browser, which would be predicted to have fewer scratches due to eating less abrasive materials. Candidates for ingested abrasives include silica-rich plants like horsetails and soil that was accidentally ingested due to feeding at ground level. The tooth structure indicates combined slicing and grinding capabilities.
Reports of gastroliths, or stomach stones, in the hadrosaurid Claosaurus are actually based on a probable double misidentification. First, the specimen is actually of Edmontosaurus annectens. Barnum Brown, who discovered the specimen in 1900, referred to it as Claosaurus because E. annectens was thought to be a species of Claosaurus at the time. Additionally, it is more likely that the supposed gastroliths represent gravel washed in during burial.
Locomotion

Like other hadrosaurids, Edmontosaurus is thought to have been a facultative biped, meaning that it mostly moved on four legs, but could adopt a bipedal stance when needed. It probably went on all fours when standing still or moving slowly, and switched to using the hind legs alone when moving more rapidly. Research conducted by computer modeling in 2007 suggests that Edmontosaurus could run at high speeds, perhaps up to 45 kilometres per hour (28 mph). Further simulations using a subadult specimen estimated as weighing 715 kilograms (1,576 lb) when alive produced a model that could run or hop bipedally, use a trot, pace, or single foot symmetric quadrupedal gait, or move at a gallop. The researchers found to their surprise that the fastest gait was kangaroo-like hopping (maximum simulated speed of 17.3 metres per second (62 km/h; 39 mph)), which they regarded as unlikely based on the size of the animal and lack of hopping footprints in the fossil record, and instead interpreted the result as indicative of an inaccuracy in their simulation. The fastest non-hopping gaits were galloping (maximum simulated speed of 15.7 metres per second (57 km/h; 35 mph)) and running bipedally (maximum simulated speed of 14.0 metres per second (50 km/h; 31 mph)). They found weak support for bipedal running as the most likely option for high-speed movement, but did not rule out high-speed quadrupedal movement.
Interactions with theropods

The time span and geographic range of Edmontosaurus overlapped with Tyrannosaurus, and an adult specimen of E. annectens on display in the Denver Museum of Nature and Science shows evidence of a theropod bite in the tail. Counting back from the hip, the thirteenth to seventeenth vertebrae have damaged spines consistent with an attack from the right rear of the animal. One spine has a portion sheared away, and the others are kinked; three have apparent tooth puncture marks. The top of the tail was at least 2.9 metres (9.5 ft) high, and the only theropod species known from the same rock formation that was tall enough to make such an attack is T. rex.
The bones are partially healed, but the edmontosaur died before the traces of damage were completely obliterated. The damage also shows signs of bone infection. Kenneth Carpenter, who studied the specimen, noted that there also seems to be a healed fracture in the left hip which predated the attack because it was more fully healed. He suggested that the edmontosaur was a target because it may have been limping from this earlier injury. Because it survived the attack, Carpenter suggested that it may have outmaneuvered or outrun its attacker, or that the damage to its tail was incurred by the hadrosaurid using it as a weapon against the tyrannosaur.
Images for kids
-

Skeletal restoration of the E. annectens (then Claosaurus) holotype, by Othniel Charles Marsh
-

Outdated E. annectens skeleton mounts, formerly referred to Anatosaurus
-

Skin impression of the specimen nicknamed "Dakota", which was found in 1999
-

Mounted skeletons of a juvenile and adult E. annectens, Houston Museum of Natural Science
-
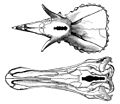
A 1905 chart showing the relatively small brains of a Triceratops (top) and E. annectens
-

E. annectens skull preserving the keratinous beak (partially removed on the right side by accident), Natural History Museum of Los Angeles
-

Horseshoe Canyon Formation near Drumheller. The dark bands are coal seams.
-

Fossils of Edmontosaurus discovered in eastern Montana (Hell Creek Formation).
-
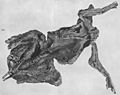
Possible gut contents were reported from the "Trachodon mummy" at the American Museum of Natural History, but were never described.
-

The Senckenberg Museum specimen
-

Edmontosaurus annectens mounted as being hunted by a Tyrannosaurus, which would have preyed on the species in life











See also
 In Spanish: Edmontosaurus para niños
In Spanish: Edmontosaurus para niños
